Glycine (ECMDB00123) (M2MDB000046)
| Record Information | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Version | 2.0 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Creation Date | 2012-05-31 09:57:49 -0600 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Update Date | 2015-09-13 12:56:06 -0600 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Secondary Accession Numbers |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Identification | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Name: | Glycine | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Description | Glycine is a simple amino acid. The glycine cleavage enzyme system comprises four proteins: P-, T-, H- and L-proteins (EC 1.4.4.2, EC 2.1.2.10 and EC 1.8.1.4 for P-, T- and L-proteins). The glycine cleavage system catalyses the oxidative conversion of glycine into carbon dioxide and ammonia, with the remaining one-carbon unit transferred to folate as methylenetetrahydrofolate. It is the main catabolic pathway for glycine and it also contributes to one-carbon metabolism. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Structure | 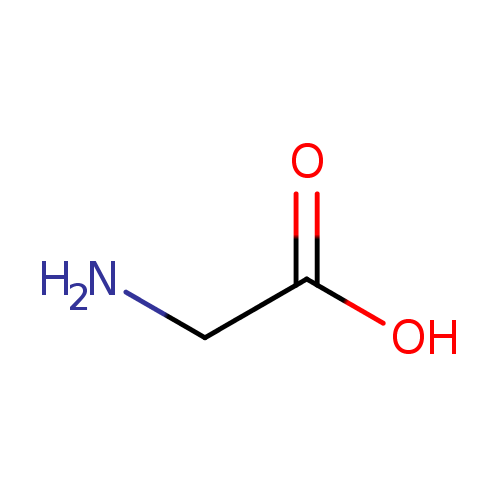 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Synonyms: |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Chemical Formula: | C2H5NO2 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Weight: | Average: 75.0666 Monoisotopic: 75.032028409 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| InChI Key: | DHMQDGOQFOQNFH-UHFFFAOYSA-N | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| InChI: | InChI=1S/C2H5NO2/c3-1-2(4)5/h1,3H2,(H,4,5) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| CAS number: | 56-40-6 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| IUPAC Name: | 2-aminoacetic acid | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Traditional IUPAC Name: | glycine | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| SMILES: | NCC(O)=O | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Chemical Taxonomy | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Description | belongs to the class of organic compounds known as alpha amino acids. These are amino acids in which the amino group is attached to the carbon atom immediately adjacent to the carboxylate group (alpha carbon). | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Kingdom | Organic compounds | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Super Class | Organic acids and derivatives | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Class | Carboxylic acids and derivatives | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Sub Class | Amino acids, peptides, and analogues | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Direct Parent | Alpha amino acids | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Alternative Parents | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Substituents |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Molecular Framework | Aliphatic acyclic compounds | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| External Descriptors |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Physical Properties | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| State: | Solid | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Charge: | 0 | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Melting point: | 504 deg F | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experimental Properties: |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Predicted Properties |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Biological Properties | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Cellular Locations: | Cytoplasm | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Reactions: | Cysteinylglycine + Water > L-Cysteine + Glycine L-Threonine <> Acetaldehyde + Glycine L-Allothreonine > Acetaldehyde + Glycine Glycine + NAD + Tetrahydrofolic acid > Carbon dioxide + 5,10-Methylene-THF + NADH + Ammonium Adenosine triphosphate + Glycine + tRNA(Gly) + tRNA(Gly) <> Adenosine monophosphate + Glycyl-tRNA(Gly) + Pyrophosphate + Glycyl-tRNA(Gly) Water + L-Prolinylglycine > Glycine + L-Proline Water + Oxygen + Sarcosine > Formaldehyde + Glycine + Hydrogen peroxide L-Serine + Tetrahydrofolic acid <> Glycine + Water + 5,10-Methylene-THF Adenosine triphosphate + gamma-Glutamylcysteine + Glycine <> ADP + Glutathione + Hydrogen ion + Phosphate Acetyl-CoA + Glycine <> L-2-Amino-3-oxobutanoic acid + Coenzyme A Adenosine triphosphate + Glycine + 5-Phosphoribosylamine <> ADP + Glycineamideribotide + Hydrogen ion + Phosphate Adenosine triphosphate + gamma-Glutamylcysteine + Glycine <> ADP + Phosphate + Glutathione Cysteinylglycine + Water <> L-Cysteine + Glycine 5,10-Methylene-THF + Glycine + Water <> Tetrahydrofolic acid + L-Serine Glycine + Tetrahydrofolic acid + NAD <> 5,10-Methylene-THF + Ammonia + Carbon dioxide + NADH + Hydrogen ion Glycine + Lipoylprotein <> S-Aminomethyldihydrolipoylprotein + Carbon dioxide Adenosine triphosphate + Glycine + tRNA(Gly) <> Adenosine monophosphate + Pyrophosphate + Glycyl-tRNA(Gly) Adenosine triphosphate + 5-Phosphoribosylamine + Glycine <> ADP + Phosphate + Glycineamideribotide R-S-Cysteinylglycine + Water <> S-Substituted L-cysteine + Glycine L-Allothreonine <> Glycine + Acetaldehyde L-Serine + 5,6,7,8-Tetrahydromethanopterin <> 5,10-Methylenetetrahydromethanopterin + Glycine + Water NAD + Glycine + Tetrahydrofolic acid > Hydrogen ion + 5,10-Methylene-THF + Ammonia + Carbon dioxide + NADH Glycine + Acetyl-CoA <> Hydrogen ion + L-2-Amino-3-oxobutanoic acid + Coenzyme A Glycine + gamma-Glutamylcysteine + Adenosine triphosphate > Hydrogen ion + Glutathione + Phosphate + ADP L-<i>threo</i>-3-phenylserine benzaldehyde + Glycine DL-allothreonine <> Acetaldehyde + Glycine More...4-Hydroxy-L-threonine <> Glycolaldehyde + Glycine gly-met + Water > Glycine + L-Methionine ala-gly + Water > L-Alanine + Glycine gly-asn + Water > Glycine + L-Asparagine gly-gln + Water > Glycine + L-Glutamine glycyl-L-glutamate + Water > Glycine + L-Glutamate gly-asp + Water > Glycine + L-Aspartic acid glycylproline + Water > Glycine + L-Proline L-Threonine > Acetaldehyde + Glycine Glycine + H-protein-lipoyllysine > H-protein-S-aminomethyldihydrolipoyllysine + Carbon dioxide 5,10-Methylene-THF + Glycine + Water > Tetrahydrofolic acid + L-Serine Adenosine triphosphate + gamma-Glutamylcysteine + Glycine > ADP + Inorganic phosphate + Glutathione Acetyl-CoA + Glycine > CoA + L-2-Amino-3-oxobutanoic acid Adenosine triphosphate + 5-Phosphoribosylamine + Glycine > ADP + Inorganic phosphate + 5'-Phospho-ribosylglycinamide Adenosine triphosphate + Glycine + tRNA(Gly) > Adenosine monophosphate + Pyrophosphate + glycyl-tRNA(Gly) Tetrahydrofolic acid + L-Serine + Tetrahydrofolic acid + L-Serine <> 5,10-Methylene-THF + Glycine + Water + 5,10-Methylene-THF L-Alanine + Glyoxylic acid + L-Alanine <> Glycine + Pyruvic acid Glycine + Adenosine triphosphate + Hydrogen ion + tRNA(gly) > Adenosine monophosphate + Pyrophosphate + Glycyl-tRNA(Gly) gamma-Glutamylcysteine + Glycine + Adenosine triphosphate > Hydrogen ion + Phosphate + Adenosine diphosphate + Glutathione + ADP 5-Phosphoribosylamine + Glycine + Adenosine triphosphate + 5-Phosphoribosylamine > N1-(5-phospho-β-D-ribosyl)glycinamide + Phosphate + Adenosine diphosphate + Hydrogen ion + ADP L-2-Amino-3-oxobutanoic acid + Coenzyme A > Acetyl-CoA + Glycine Adenosine triphosphate + Glycine + tRNA(Gly) <> Adenosine monophosphate + Glycyl-tRNA(Gly) + Pyrophosphate Adenosine triphosphate + gamma-Glutamylcysteine + Glycine <> ADP + Glutathione + Hydrogen ion + Phosphate L-Threonine <> Acetaldehyde + Glycine L-Allothreonine > Acetaldehyde + Glycine Cysteinylglycine + Water > L-Cysteine + Glycine Adenosine triphosphate + Glycine + 5 5-Phosphoribosylamine <> ADP + Glycineamideribotide + Hydrogen ion + Phosphate Glycine + Lipoylprotein <> S-Aminomethyldihydrolipoylprotein + Carbon dioxide Glycine + Tetrahydrofolic acid + NAD <>5 5,10-Methylene-THF + Ammonia + Carbon dioxide + NADH + Hydrogen ion Adenosine triphosphate + Glycine + tRNA(Gly) <> Adenosine monophosphate + Glycyl-tRNA(Gly) + Pyrophosphate L-Allothreonine > Acetaldehyde + Glycine Glycine + Tetrahydrofolic acid + NAD <>5 5,10-Methylene-THF + Ammonia + Carbon dioxide + NADH + Hydrogen ion | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| SMPDB Pathways: |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| KEGG Pathways: |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| EcoCyc Pathways: |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Concentrations | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Find out more about how we convert literature concentrations. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Spectra | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Spectra: | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| References | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| References: |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Synthesis Reference: | Anslow, Winston K.; King, Harold. Synthesis of glycine. Journal of the Chemical Society (1929), 2163-6. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Material Safety Data Sheet (MSDS) | Download (PDF) | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Links | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| External Links: |
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

Enzymes
- General function:
- Involved in nucleotide binding
- Specific function:
- ATP + glycine + tRNA(Gly) = AMP + diphosphate + glycyl-tRNA(Gly)
- Gene Name:
- glyQ
- Uniprot ID:
- P00960
- Molecular weight:
- 34774
Reactions
| ATP + glycine + tRNA(Gly) = AMP + diphosphate + glycyl-tRNA(Gly). |
- General function:
- Involved in arginine-tRNA ligase activity
- Specific function:
- ATP + glycine + tRNA(Gly) = AMP + diphosphate + glycyl-tRNA(Gly)
- Gene Name:
- glyS
- Uniprot ID:
- P00961
- Molecular weight:
- 76812
Reactions
| ATP + glycine + tRNA(Gly) = AMP + diphosphate + glycyl-tRNA(Gly). |
- General function:
- Involved in ATP binding
- Specific function:
- ATP + gamma-L-glutamyl-L-cysteine + glycine = ADP + phosphate + glutathione
- Gene Name:
- gshB
- Uniprot ID:
- P04425
- Molecular weight:
- 35561
Reactions
| ATP + gamma-L-glutamyl-L-cysteine + glycine = ADP + phosphate + glutathione. |
- General function:
- Involved in proteolysis
- Specific function:
- Aminopeptidase N is involved in the degradation of intracellular peptides generated by protein breakdown during normal growth as well as in response to nutrient starvation
- Gene Name:
- pepN
- Uniprot ID:
- P04825
- Molecular weight:
- 98918
Reactions
| Release of an N-terminal amino acid, Xaa-|-Yaa- from a peptide, amide or arylamide. Xaa is preferably Ala, but may be most amino acids including Pro (slow action). When a terminal hydrophobic residue is followed by a prolyl residue, the two may be released as an intact Xaa-Pro dipeptide. |
- General function:
- Involved in catalytic activity
- Specific function:
- Interconversion of serine and glycine
- Gene Name:
- glyA
- Uniprot ID:
- P0A825
- Molecular weight:
- 45316
Reactions
| 5,10-methylenetetrahydrofolate + glycine + H(2)O = tetrahydrofolate + L-serine. |
- General function:
- Involved in oxidoreductase activity
- Specific function:
- Lipoamide dehydrogenase is a component of the glycine cleavage system as well as of the alpha-ketoacid dehydrogenase complexes
- Gene Name:
- lpdA
- Uniprot ID:
- P0A9P0
- Molecular weight:
- 50688
Reactions
| Protein N(6)-(dihydrolipoyl)lysine + NAD(+) = protein N(6)-(lipoyl)lysine + NADH. |
- General function:
- Involved in glycine C-acetyltransferase activity
- Specific function:
- Catalyzes the cleavage of 2-amino-3-ketobutyrate to glycine and acetyl-CoA
- Gene Name:
- kbl
- Uniprot ID:
- P0AB77
- Molecular weight:
- 43117
Reactions
| Acetyl-CoA + glycine = CoA + 2-amino-3-oxobutanoate. |
- General function:
- Involved in hydrolase activity
- Specific function:
- Dipeptidase with broad substrate specificity. Requires dipeptide substrates with an unblocked N-terminus and the amino group in the alpha or beta position. Non-protein amino acids and proline are not accepted in the C-terminal position, whereas some dipeptide amides and formyl amino acids are hydrolyzed. Also shows cysteinylglycinase activity, which is sufficient for E.coli to utilize cysteinylglycine as a cysteine source
- Gene Name:
- pepD
- Uniprot ID:
- P15288
- Molecular weight:
- 52915
Reactions
| Hydrolysis of dipeptides, preferentially hydrophobic dipeptides including prolyl amino acids. |
- General function:
- Involved in ATP binding
- Specific function:
- ATP + 5-phospho-D-ribosylamine + glycine = ADP + phosphate + N(1)-(5-phospho-D-ribosyl)glycinamide
- Gene Name:
- purD
- Uniprot ID:
- P15640
- Molecular weight:
- 45940
Reactions
| ATP + 5-phospho-D-ribosylamine + glycine = ADP + phosphate + N(1)-(5-phospho-D-ribosyl)glycinamide. |
- General function:
- Involved in aminomethyltransferase activity
- Specific function:
- The glycine cleavage system catalyzes the degradation of glycine
- Gene Name:
- gcvT
- Uniprot ID:
- P27248
- Molecular weight:
- 40146
Reactions
| [Protein]-S(8)-aminomethyldihydrolipoyllysine + tetrahydrofolate = [protein]-dihydrolipoyllysine + 5,10-methylenetetrahydrofolate + NH(3). |
- General function:
- Involved in glycine dehydrogenase (decarboxylating) activity
- Specific function:
- The glycine cleavage system catalyzes the degradation of glycine. The P protein binds the alpha-amino group of glycine through its pyridoxal phosphate cofactor; CO(2) is released and the remaining methylamine moiety is then transferred to the lipoamide cofactor of the H protein
- Gene Name:
- gcvP
- Uniprot ID:
- P33195
- Molecular weight:
- 104376
Reactions
| Glycine + H-protein-lipoyllysine = H-protein-S-aminomethyldihydrolipoyllysine + CO(2). |
- General function:
- Involved in aminopeptidase activity
- Specific function:
- Probably plays an important role in intracellular peptide degradation
- Gene Name:
- pepB
- Uniprot ID:
- P37095
- Molecular weight:
- 46180
Reactions
| Release of an N-terminal amino acid, Xaa, from a peptide or arylamide. Xaa is preferably Glu or Asp but may be other amino acids, including Leu, Met, His, Cys and Gln. |
- General function:
- Involved in aminopeptidase activity
- Specific function:
- Presumably involved in the processing and regular turnover of intracellular proteins. Catalyzes the removal of unsubstituted N-terminal amino acids from various peptides. Required for plasmid ColE1 site-specific recombination but not in its aminopeptidase activity. Could act as a structural component of the putative nucleoprotein complex in which the Xer recombination reaction takes place
- Gene Name:
- pepA
- Uniprot ID:
- P68767
- Molecular weight:
- 54879
Reactions
| Release of an N-terminal amino acid, Xaa-|-Yaa-, in which Xaa is preferably Leu, but may be other amino acids including Pro although not Arg or Lys, and Yaa may be Pro. Amino acid amides and methyl esters are also readily hydrolyzed, but rates on arylamides are exceedingly low. |
| Release of an N-terminal amino acid, preferentially leucine, but not glutamic or aspartic acids. |
- General function:
- Involved in lyase activity
- Specific function:
- Catalyzes the cleavage of L-allo-threonine and L- threonine to glycine and acetaldehyde. L-threo-phenylserine and L- erythro-phenylserine are also good substrates
- Gene Name:
- ltaE
- Uniprot ID:
- P75823
- Molecular weight:
- 36494
Reactions
| L-threonine = glycine + acetaldehyde. |
| L-allo-threonine = glycine + acetaldehyde. |
- General function:
- Involved in oxidoreductase activity
- Specific function:
- Catalyzes the oxidative demethylation of N-methyl-L- tryptophan. Can also use other N-methyl amino acids, including sarcosine, which, however, is a poor substrate
- Gene Name:
- solA
- Uniprot ID:
- P40874
- Molecular weight:
- 40902
Reactions
| N-methyl-L-tryptophan + H(2)O + O(2) = L-tryptophan + formaldehyde + H(2)O(2). |
- General function:
- Amino acid transport and metabolism
- Specific function:
- The glycine cleavage system catalyzes the degradation of glycine. The H protein shuttles the methylamine group of glycine from the P protein to the T protein
- Gene Name:
- gcvH
- Uniprot ID:
- P0A6T9
- Molecular weight:
- 13811
Transporters
- General function:
- Involved in proteolysis
- Specific function:
- Aminopeptidase N is involved in the degradation of intracellular peptides generated by protein breakdown during normal growth as well as in response to nutrient starvation
- Gene Name:
- pepN
- Uniprot ID:
- P04825
- Molecular weight:
- 98918
Reactions
| Release of an N-terminal amino acid, Xaa-|-Yaa- from a peptide, amide or arylamide. Xaa is preferably Ala, but may be most amino acids including Pro (slow action). When a terminal hydrophobic residue is followed by a prolyl residue, the two may be released as an intact Xaa-Pro dipeptide. |
- General function:
- Involved in sodium:amino acid symporter activity
- Specific function:
- Specific function unknown
- Gene Name:
- yaaJ
- Uniprot ID:
- P30143
- Molecular weight:
- 51662
- General function:
- Involved in nucleotide binding
- Specific function:
- Probably part of a binding-protein-dependent transport system yecCS for an amino acid. Probably responsible for energy coupling to the transport system
- Gene Name:
- yecC
- Uniprot ID:
- P37774
- Molecular weight:
- 27677
- General function:
- Involved in transporter activity
- Specific function:
- Probably part of the binding-protein-dependent transport system yecCS for an amino acid; probably responsible for the translocation of the substrate across the membrane
- Gene Name:
- yecS
- Uniprot ID:
- P0AFT2
- Molecular weight:
- 24801
- General function:
- Involved in transport
- Specific function:
- Permease that is involved in the transport across the cytoplasmic membrane of D-alanine, D-serine and glycine
- Gene Name:
- cycA
- Uniprot ID:
- P0AAE0
- Molecular weight:
- 51659
- General function:
- Involved in transporter activity
- Specific function:
- Probable transporter whose substrate is unknown. Is not involved in aerobic D-malate transport
- Gene Name:
- yeaV
- Uniprot ID:
- P0ABD1
- Molecular weight:
- 52881
- General function:
- Involved in transporter activity
- Specific function:
- Non-specific porin
- Gene Name:
- ompN
- Uniprot ID:
- P77747
- Molecular weight:
- 41220
- General function:
- Involved in transporter activity
- Specific function:
- Uptake of inorganic phosphate, phosphorylated compounds, and some other negatively charged solutes
- Gene Name:
- phoE
- Uniprot ID:
- P02932
- Molecular weight:
- 38922
- General function:
- Involved in transporter activity
- Specific function:
- OmpF is a porin that forms passive diffusion pores which allow small molecular weight hydrophilic materials across the outer membrane. It is also a receptor for the bacteriophage T2
- Gene Name:
- ompF
- Uniprot ID:
- P02931
- Molecular weight:
- 39333
- General function:
- Involved in transporter activity
- Specific function:
- Forms passive diffusion pores which allow small molecular weight hydrophilic materials across the outer membrane
- Gene Name:
- ompC
- Uniprot ID:
- P06996
- Molecular weight:
- 40368